Lisaks genotüübile on olemas modifikatsiooni varieeruvus , mida peetakse vastuseks keskkonnatingimuste muutustele ja mida täheldatakse seni, kuni neid muutusi põhjustav tegur toimib. Modifikatsiooni varieeruvus(seda nimetatakse ka fenotüüpne varieeruvus) avaldub fenotüübi tasemel ega mõjuta genotüüpi.
Fenotüüpne varieeruvus avaldub valdavas enamuses elanikkonnast, samas kui mutatsioonide varieeruvuse korral toimub genotüübi muutus ainult üksikrakkudes.
Modifikatsioon on raku ainevahetuse plastilisuse tulemus, mis viib "vaikivate" geenide fenotüübilise avaldumiseni konkreetsetes tingimustes. Seega modifikatsioonimuutused toimuvad muutumatu raku genotüübi raames.
Muudatuste muutmisel on mitmeid ilminguid. Kõige kuulsam adaptiivsed modifikatsioonid , st. ... mittepärilikud muutused, mis on organismile kasulikud ja aitavad kaasa selle säilimisele muutunud tingimustes.
Adaptiivsete modifikatsioonide põhjused peituvad geenitoime reguleerimise mehhanismides. Selle näiteks on bakterirakkude kohanemine E. coli laktoosile uueks substraadiks: nendes tingimustes hakkavad sünteesima indutseeritavad ensüümid, st geenide fenotüüpne avaldumine "vaikne" laktoosi puudumisel söötmes.
On leitud hulk baktereid universaalne adaptiivne reaktsioon vastuseks erinevatele pingelistele mõjudele (kõrged ja madalad temperatuurid, järsk pH nihe jne). Sellisel juhul avaldub adaptiivne reaktsioon väikese sarnaste valkude rühma, mida nimetatakse, intensiivseks sünteesiks kuumašoki valgud ja nähtus - kuumašoki sündroom ... Bakteriraku stressirohke mõju all pärsitakse selles tavaliste valkude sünteesi, kuid indutseeritakse väikese proteiinirühma süntees, mille ülesandeks on stressi vastu võitlemine, kaitstes kõige olulisemaid rakustruktuure, eelkõige nukleoide ja membraanid. Arvatakse, et adaptiivsed modifikatsioonid suurendavad keha võimet ellu jääda ja paljuneda laiemates keskkonnatingimustes. Sellest tulenevad muudatused võivad olla suhteliselt stabiilne, need võivad kesta mitu põlvkonda või vastupidi väga labiilne.
Siiski ei saa kõiki muudatusi kohandada. Intensiivsel kokkupuutel paljude ainetega, päritud muutused, juhuslikud nende tekitanud mõju suhtes. Selliste fenotüüpiliselt muutunud rakkude ilmumise põhjused on seotud nende ainete põhjustatud vigadega tõlkeprotsessis.
Kohanduvate muudatuste tähendus:
- anda teatud panus evolutsiooniprotsessi;
- laiendada keha võimet ellu jääda ja paljuneda laiemates keskkonnatingimustes. Nendes tingimustes tekkivaid pärilikke muutusi kiirendab looduslik valik ja sel viisil areneb aktiivsemalt uusi ökoloogilisi nišše ja saavutatakse nendega tõhusam kohanemisvõime.
Bakterid on vanim organismide rühm, mis praegu Maal eksisteerib. Esimesed bakterid ilmusid ilmselt rohkem kui 3,5 miljardit aastat tagasi ja peaaegu miljard aastat olid meie planeedil ainsad elusolendid. Kuna need olid eluslooduse esimesed esindajad, oli nende kehal ürgne struktuur.
Aja jooksul on nende struktuur muutunud keerulisemaks, kuid tänapäevani peetakse baktereid kõige primitiivsemateks üherakulisteks organismideks. Huvitav on see, et mõned bakterid säilitavad endiselt oma iidsete esivanemate ürgsed tunnused. Seda täheldatakse bakterites, mis elavad kuumades väävliallikates ja veehoidlate põhjas asuvatel anoksilistel muldadel.
Enamik baktereid on värvitu. Vaid vähesed on lillad või rohelised. Kuid paljude bakterite kolooniatel on ere värv, mis on tingitud värvilise aine keskkonda sattumisest või rakkude pigmentatsioonist.
Bakterimaailma pioneeriks oli 17. sajandi Hollandi loodusteadlane Anthony Leeuwenhoek, kes lõi esimesena täiusliku suurendusklaasi mikroskoobi, mis suurendab objekte 160–270 korda.
Bakterid klassifitseeritakse prokarüootideks ja eraldatakse eraldi kuningriigiks - bakteriteks.
Bakterid on arvukad ja mitmekesised organismid. Need on erineva kujuga.
| Bakterite nimi | Bakterite kuju | Pilt bakteritest |
| Cocci | Sfääriline | |
| Bacillus |  | Vardakujuline |
| Vibrio | Koma kaardus | |
| Spirillum |  | Spiraal |
| Streptokokid |  | Cocci kett |
| Stafülokokid |  | Kookide kobarad |
| Diplokokid | Kaks ümmargust bakterit, mis on suletud ühte limaskesta kapslisse |
Bakterite hulgas on liikuvad ja liikumatud vormid. Mobiilsed liiguvad lainekujuliste kokkutõmmete tõttu või flagella (keerdunud spiraalsed kiud) abil, mis koosnevad spetsiaalsest flagelliinvalgust. Flageleid võib olla üks või mitu. Need asuvad mõnes bakteris raku ühes otsas, teistes - kahel või kogu pinnal.

Kuid liikumine on omane paljudele teistele bakteritele, millel puuduvad lipukesed. Niisiis, väljastpoolt limaga kaetud bakterid on võimelised liikuma.
Mõnel vee- ja mullabakteril, millel puuduvad lipukesed, on tsütoplasmas gaasivakuoolid. Rakus võib olla 40–60 vakuooli. Igaüks neist on täidetud gaasiga (arvatavasti lämmastikuga). Reguleerides gaasikogust vakuoolides, võivad vesibakterid sukelduda veesambasse või tõusta selle pinnale ning mullabakterid võivad liikuda mulla kapillaarides.
Organisatsiooni lihtsuse ja tagasihoidlikkuse tõttu on bakterid looduses laialt levinud. Baktereid leidub kõikjal: isegi kõige puhtama allikavee tilgas, mulla terades, õhus, kivimitel, polaarlumes, kõrbeliival, ookeanipõhjas, suurest sügavusest ekstraheeritud õlis ja isegi kuumaveeallikates temperatuuril umbes 80 ° C. Nad elavad taimedel, puuviljadel, erinevatel loomadel ja inimestel soolestikus, suus, jäsemetel, keha pinnal.
Bakterid on kõige väiksemad ja arvukamad elusolendid. Väikese suuruse tõttu tungivad nad kergesti kõikidesse pragudesse, pragudesse, pooridesse. Nad on väga vastupidavad ja kohandatud erinevate eksistentsitingimustega. Nad taluvad kuivatamist, tugevat külma, kuumutamist kuni 90 ° C, kaotamata oma elujõulisust.
Maal pole praktiliselt ühtegi kohta, kus baktereid ei leiduks, kuid erinevates kogustes. Bakterite elutingimused on mitmekesised. Üks neist vajab hapnikku õhus, teised seda ei vaja ja suudavad elada hapnikuvabas keskkonnas.
Õhus: bakterid tõusevad atmosfääri ülemisse ossa kuni 30 km. ja veel.
Eriti palju on neid mullas. Üks aasta mulda võib sisaldada sadu miljoneid baktereid.
Vees: pinnaveekihtides avatud reservuaarides. Kasulikud vesibakterid mineraliseerivad orgaanilisi jääke.
Elusorganismides: patogeensed bakterid sisenevad kehasse väliskeskkonnast, kuid ainult soodsates tingimustes põhjustavad haigusi. Sümbiootik elab seedeorganites, aidates toitu lagundada ja assimileerida, sünteesida vitamiine.
Bakterirakk on riietatud spetsiaalsesse tihedasse kesta - rakuseina, mis täidab kaitse- ja tugifunktsioone ning annab ka bakteritele püsiva iseloomuliku kuju. Bakteri rakusein meenutab taimeraku membraani. See on läbilaskev: selle kaudu pääsevad toitained vabalt rakku ja ainevahetusproduktid väljuvad keskkonda. Sageli tekitavad bakterid rakuseina peale täiendava kaitsva lima kihi - kapsli. Kapsli paksus võib olla mitu korda suurem kui lahtri enda läbimõõt, kuid see võib olla väga väike. Kapsel ei ole raku kohustuslik osa; see moodustub sõltuvalt bakterite sisenemise tingimustest. See hoiab ära bakterite kuivamise.
Mõnede bakterite pinnal on pikad lipukesed (üks, kaks või palju) või lühikesed õhukesed villid. Flagella pikkus võib olla mitu korda suurem kui bakterite keha mõõtmed. Flagella ja villide abil liiguvad bakterid.

Bakteriraku sees on tihe, liikumatu tsütoplasma. Sellel on kihiline struktuur, puuduvad vakuoolid, seetõttu asuvad tsütoplasma aines mitmesugused valgud (ensüümid) ja reservtoitained. Bakterirakkudel puudub tuum. Nende rakkude keskosas on aine, mis kannab pärilikku teavet. Bakterid, - nukleiinhape - DNA. Kuid see aine ei moodustu tuumaks.
Bakteriraku sisemine korraldus on keeruline ja sellel on oma eripärad. Tsütoplasma eraldab rakuseinast tsütoplasmaatiline membraan. Tsütoplasmas eristatakse põhiainet ehk maatriksit, ribosoome ja väikest hulka membraani struktuure, mis täidavad erinevaid funktsioone (mitokondrite analoogid, endoplasmaatiline retikulum, Golgi aparaat). Bakterirakkude tsütoplasmas on sageli erineva kuju ja suurusega graanuleid. Graanulid võivad koosneda ühenditest, mis toimivad energia ja süsiniku allikana. Rasvatilku leidub ka bakterirakus.
Raku keskosas paikneb tuumaine - DNA, mis ei ole tsütoplasmast membraaniga piiritletud. See on tuuma analoog - nukleoid. Nukleoidil pole membraani, tuuma ja kromosoomide komplekti.
Bakteritel on erinevad toitumisviisid. Nende hulka kuuluvad autotroofid ja heterotroofid. Autotroofid on organismid, mis võivad iseseisvalt moodustada oma toitumiseks orgaanilisi aineid.
Taimed vajavad lämmastikku, kuid nad ise ei saa õhust lämmastikku omastada. Mõned bakterid ühendavad õhus olevad lämmastiku molekulid teiste molekulidega, et muuta taimedele kättesaadavaks ained.
Need bakterid asetuvad noorte juurte rakkudesse, mis põhjustab juurtel paksenemist, mida nimetatakse sõlmedeks. Sellised sõlmed moodustuvad kaunviljade perekonna taimede juurtel ja mõnedel teistel taimedel.

Juured varustavad baktereid süsivesikutega ja bakterid annavad juurtele lämmastikku sisaldavaid aineid, mida taim suudab omastada. Nende kooselu on vastastikku kasulik.
Taimede juured eritavad palju orgaanilisi aineid (suhkruid, aminohappeid ja teisi), millest bakterid toituvad. Seetõttu asustub juure ümbritsevasse mullakihti eriti suur hulk baktereid. Need bakterid muudavad surnud taimejäägid taimele kättesaadavaks aineks. Seda mullakihti nimetatakse risosfääriks.
Sõlmebakterite juurekoesse tungimise kohta on mitmeid hüpoteese:
Sõlmebakterite juurkoesse viimise protsess koosneb kahest etapist:
Enamasti paljunev rakk paljuneb aktiivselt, moodustab nn nakkuslikud kiud ja juba selliste hõõgniitide kujul liigub taimekoesse. Nakkusniidist vabanenud sõlmebakterid jätkavad peremeeskoes paljunemist.

Taimerakud, mis on täidetud kiiresti paljunevate sõlmebakterite rakkudega, hakkavad kiiresti jagunema. Noore sõlme ühendamine liblikõielise taime juurega toimub tänu veresoonte-kiudude kimpudele. Toimimisperioodil on sõlmed tavaliselt tihedad. Optimaalse aktiivsuse ilmnemise ajaks omandavad sõlmed roosa värvi (pigmendi leghemoglobiini tõttu). Ainult need bakterid, mis sisaldavad leghemoglobiini, on võimelised lämmastikku siduma.
Sõlmebakterid loovad mulla hektari kohta kümneid ja sadu kilogramme lämmastikväetisi.
Bakterid erinevad üksteisest ainevahetuse poolest. Mõnel läheb see hapniku osalusel, teistel - ilma selle osaluseta.
Enamik baktereid toitub valmis orgaanilisest ainest. Ainult vähesed neist (sinakasrohelised või tsüanobakterid) on võimelised looma anorgaanilistest ainetest orgaanilisi aineid. Neil oli oluline roll hapniku kogunemisel Maa atmosfääri.
Bakterid neelavad aineid väljastpoolt, rebivad nende molekulid laiali, nendest osadest koguvad nad oma kesta ja täiendavad nende sisu (nii nad kasvavad) ning tarbetud molekulid visatakse välja. Bakteri kest ja membraan võimaldavad tal imada ainult vajalikke aineid.

Kui bakterite kest ja membraan oleksid täiesti mitteläbilaskvad, ei satuks rakku aineid. Kui need oleksid kõikidele ainetele läbilaskvad, seguneks raku sisu keskkonnaga - lahusega, milles bakter elab. Bakterite ellujäämiseks on vaja kesta, mis laseb läbi vajalikud ained, kuid mitte mittevajalikud.
Bakter neelab läheduses olevaid toitaineid. Mis järgmisena juhtub? Kui see suudab iseseisvalt liikuda (lipukest liigutades või lima tagasi lükates), siis liigub see seni, kuni leiab vajalikud ained.
Kui see ei saa liikuda, siis ta ootab, kuni difusioon (ühe aine molekulide võime tungida teise aine molekulide keskele) toob sinna vajalikud molekulid.
Bakterid teevad koos teiste mikroorganismide rühmadega tohutult palju keemilist tööd. Erinevaid ühendeid muundades saavad nad oma eluks vajaliku energia ja toitained. Ainevahetusprotsessid, energia hankimise meetodid ja ehitusmaterjalide vajadus nende kehas on bakterites erinevad.
Teised bakterid vastavad kõikidele süsiniku nõuetele, mis on vajalikud orgaaniliste ainete sünteesiks organismis anorgaaniliste ühendite arvelt. Neid nimetatakse autotroofideks. Autotroofsed bakterid on võimelised sünteesima orgaanilisi aineid anorgaanilistest. Nende hulgas eristatakse:
Kiirgusenergia kasutamine on kõige olulisem, kuid mitte ainus viis orgaanilise aine loomiseks süsinikdioksiidist ja veest. On teada, et bakterid ei kasuta selliseks sünteesiks energiaallikana päikesevalgust, vaid teatud anorgaaniliste ühendite - vesiniksulfiid, väävel, ammoniaak, vesinik, lämmastikhape, raudühendid - oksüdeerimisel organismide rakkudes esinevate keemiliste sidemete energiat. raud ja mangaan. Nad kasutavad selle keemilise energia kasutamisel tekkinud orgaanilist ainet oma keha rakkude ehitamiseks. Seetõttu nimetatakse seda protsessi kemosünteesiks.
Kemosünteetiliste mikroorganismide kõige olulisem rühm on nitrifitseerivad bakterid. Need bakterid elavad mullas ja viivad orgaaniliste jääkide lagunemisel tekkinud ammoniaagi oksüdatsiooni lämmastikhappeks. Viimane, reageerides pinnase mineraalühenditega, muutub lämmastikhappe sooladeks. See protsess toimub kahes etapis.
Raudbakterid muudavad raudraua oksiidiks. Tekkinud raudhüdroksiid settib ja moodustab nn rabamaaki.
Mõned mikroorganismid eksisteerivad molekulaarse vesiniku oksüdeerimisel, pakkudes seeläbi autotroofset toitumisviisi.
Vesinikbakterite iseloomulik tunnus on võime minna üle heterotroofsele eluviisile, kui neid varustatakse orgaaniliste ühenditega ja vesiniku puudumisel.
Seega on kemoautotroofid tüüpilised autotroofid, kuna need sünteesivad anorgaanilistest ainetest iseseisvalt vajalikke orgaanilisi ühendeid ega võta neid teistest organismidest, nagu heterotroofid, valmis. Kemoautotroofsed bakterid erinevad fototroofsetest taimedest nende täieliku sõltumatuse tõttu valgusest kui energiaallikast.
Mõned pigmenti sisaldavad väävlibakterid (lilla, roheline), mis sisaldavad spetsiifilisi pigmente - bakterioklorofülli, on võimelised neelama päikeseenergiat, mille abil nende organismides olev vesiniksulfiid laguneb ja vabastab vesinikuaatomid, et taastada vastavad ühendid. Sellel protsessil on fotosünteesiga palju ühist ja see erineb ainult selle poolest, et lillade ja roheliste bakterite puhul on vesiniksulfiid vesiniku doonor (aeg -ajalt - karboksüülhapped) ja rohelistel taimedel vesi. Mõlemal juhul toimub vesiniku eemaldamine ja ülekandmine neeldunud päikesekiirte energia tõttu.
Seda bakterite fotosünteesi, mis toimub ilma hapniku eraldumiseta, nimetatakse fotoreduktsiooniks. Süsinikdioksiidi fotoreduktsioon on seotud vesiniku ülekandmisega mitte veest, vaid vesiniksulfiidist:
6СО 2 + 12Н 2 S + hv → С6Н 12 О 6 + 12S = 6Н 2 О
Kemosünteesi ja bakterite fotosünteesi bioloogiline tähtsus planeedil on suhteliselt väike. Ainult keemiliselt sünteesivad bakterid mängivad looduses väävlitsüklis olulist rolli. Roheliste taimede poolt väävelhappe soolade kujul imendunud väävel väheneb ja on osa valgumolekulidest. Veelgi enam, kui surnud taime- ja loomajäägid hävitavad mädanenud bakterid, eraldub väävel vesiniksulfiidi kujul, mille väävlibakterid oksüdeerivad vabaks väävliks (või väävelhappeks), mis moodustab mullas taimele kättesaadavad sulfitid. Keemia- ja fotoautotroofsed bakterid on lämmastiku ja väävli tsüklis hädavajalikud.
Eosed tekivad bakteriraku sees. Sporulatsiooni käigus läbib bakterirakk mitmeid biokeemilisi protsesse. Vaba vee kogus selles väheneb, ensümaatiline aktiivsus väheneb. See tagab eoste vastupidavuse ebasoodsatele keskkonnatingimustele (kõrge temperatuur, kõrge soolakontsentratsioon, kuivatamine jne). Sporulatsioon on iseloomulik ainult väikesele bakterite rühmale.

Eosed on bakterite elutsüklis vabatahtlikud. Eoste moodustumine algab ainult toitainete puudusest või ainevahetusproduktide kogunemisest. Eoste kujul olevad bakterid võivad pikka aega uinuda. Bakterite eosed taluvad pikaajalist keetmist ja väga pikka külmumist. Soodsate tingimuste tekkimisel eosed idanevad ja muutuvad elujõuliseks. Bakterite eosed on kohanemine ellujäämisega ebasoodsates tingimustes.
Bakterid paljunevad, jagades ühe raku kaheks. Olles saavutanud teatud suuruse, jaguneb bakter kaheks identseks bakteriks. Siis hakkab igaüks neist toitma, kasvab, jagab jne.

Pärast rakkude pikenemist moodustub järk -järgult põikivahesein ja seejärel lahknevad tütarrakud; paljudes bakterites jäävad teatud tingimustel rakud pärast jagunemist iseloomulikeks rühmadeks. Sel juhul tekivad olenevalt jaotusplaani suunast ja jagunemiste arvust erinevad kujud. Paljunemine pungade abil toimub erandkorras bakterites.
Soodsates tingimustes toimub rakkude jagunemine paljudes bakterites iga 20-30 minuti järel. Sellise kiire paljunemisega suudab ühe bakteri järglased 5 päevaga moodustada massi, mis suudab täita kõik mered ja ookeanid. Lihtne arvutus näitab, et päevas saab moodustada 72 põlvkonda (720 000 000 000 000 000 000 rakku). Kui tõlkida kaaluks - 4720 tonni. Looduses seda aga ei juhtu, kuna enamik baktereid sureb kiiresti päikesevalguse mõjul, kuivatamise, toidupuuduse, kuumutamise ajal temperatuurini 65–100 ° C, liikidevahelise võitluse tagajärjel jne.
Piisavalt toitu omastanud bakter (1) suureneb (2) ja hakkab valmistuma paljunemiseks (rakkude jagunemine). Selle DNA (bakterites on DNA molekul ringis suletud) kahekordistub (bakter toodab selle molekuli koopia). Mõlemad DNA molekulid (3,4) osutuvad bakteri seina külge kinnitatuks ja kui bakterid venivad, lähevad nad külgedele (5,6). Esmalt jagatakse nukleotiid, seejärel tsütoplasma.

Pärast kahe DNA molekuli lahknemist ilmneb bakteritele kitsendus, mis jagab bakteri keha järk -järgult kaheks osaks, millest igaüks sisaldab DNA molekuli (7).
See juhtub (heinapulgas), kaks bakterit kleepuvad kokku ja nende vahele moodustub sild (1,2).

Silla kaudu transporditakse DNA ühest bakterist teise (3). Kord ühes bakteris põimuvad DNA molekulid, kleepuvad mõnes kohas kokku (4), pärast mida vahetavad nad sektsioone (5).
Bakterid on looduses ainete üldises ringluses kõige olulisem lüli. Taimed loovad keerukaid orgaanilisi aineid süsinikdioksiidist, veest ja mulla mineraalsooladest. Need ained naasevad mulda koos surnud seente, taimede ja loomade laipadega. Bakterid lagundavad keerulised ained lihtsateks, mida taimed taas kasutavad.
Bakterid hävitavad surnud taimede ja loomade laipade keerulisi orgaanilisi aineid, elusorganismide väljaheiteid ja erinevaid jääkaineid. Neid orgaanilisi aineid toites muudavad saprofüütilised mädanenud bakterid need huumuseks. Need on meie planeedi omamoodi korraldused. Seega osalevad bakterid aktiivselt looduses olevate ainete ringis.
Kuna bakterid on peaaegu kõikjal laialt levinud ja neid leidub tohutul hulgal, määravad need suuresti looduses esinevad erinevad protsessid. Sügisel langevad puude ja põõsaste lehed, kõrreliste õhuvõrsed surevad ära, vanad oksad kukuvad maha, aeg -ajalt langevad vanade puude tüved. Kõik see muutub järk -järgult huumuseks. 1 cm 3 juures. Metsamulla pindmine kiht sisaldab sadu miljoneid mitme liigi saprofüütilisi mullabaktereid. Need bakterid muudavad huumuse erinevateks mineraalideks, mida taimejuured võivad mullast imada.
Mõned mullabakterid suudavad õhust lämmastikku imada, kasutades seda eluprotsessides. Need lämmastikku siduvad bakterid elavad iseseisvalt või asuvad kaunviljade juurtesse. Pärast kaunviljade juurte tungimist põhjustavad need bakterid juurerakkude kasvu ja neile sõlmede moodustumist.
Need bakterid eraldavad lämmastikuühendeid, mida taimed kasutavad. Bakterid saavad taimedelt süsivesikuid ja mineraalsooli. Seega on liblikõielise taime ja sõlmebakterite vahel tihe seos, mis on kasulik nii ühele kui ka teisele organismile. Seda nähtust nimetatakse sümbioosiks.
Tänu sümbioosile sõlmebakteritega rikastavad kaunviljad mulda lämmastikuga, aidates saaki suurendada.
Mikroorganismid on kõikjal. Ainsad erandid on aktiivsete vulkaanide kraatrid ja väikesed alad plahvatanud aatomipommide epitsentrites. Mikrofloora olemasolu ja arengut ei sega ei Antarktika madalad temperatuurid, geisrite keevad joad ega soolalahuste küllastunud soolalahused, mäetippude tugev insolatsioon ega tuumareaktorite tugev kiiritamine. Kõik elusolendid suhtlevad pidevalt mikroorganismidega, olles sageli mitte ainult nende hoidlad, vaid ka levitajad. Mikroorganismid on meie planeedi aborigeenid, kes assimileerivad aktiivselt kõige uskumatumaid looduslikke substraate.
Bakterite arv mullas on äärmiselt suur - sadu miljoneid ja miljardeid isendeid grammi kohta. Neid on mullas palju rohkem kui vees ja õhus. Bakterite koguarv pinnases on erinev. Bakterite arv sõltub mulla tüübist, nende seisundist, kihtide sügavusest.
Mullaosakeste pinnal paiknevad mikroorganismid väikestes mikrokolooniates (20–100 rakku kummaski). Sageli arenevad nad paksude orgaaniliste ainete hüübidena, elavate ja surevate taimejuurte, õhukeste kapillaaride ja tükkide sees.
Mulla mikrofloora on väga mitmekesine. Bakterite füsioloogilisi rühmi on erinevaid: mädanevad bakterid, nitrifitseerivad, lämmastikku fikseerivad, väävlibakterid jne. Nende hulgas on aeroobseid ja anaeroobseid, eoseid ja eoseid. Mikrofloora on üks mulla moodustumise teguritest.
Mikroorganismide arengupiirkond mullas on elavate taimede juurtega külgnev ala. Seda nimetatakse risosfääriks ja selles sisalduvate mikroorganismide kogu nimetatakse risosfääri mikroflooraks.
Vesi on looduslik keskkond, kus mikroorganisme kasvab palju. Enamik neist satub vette mullast. Tegur, mis määrab bakterite arvu vees, toitainete olemasolu selles. Kõige puhtamad on arteesiakaevud ja allikaveed. Avatud veehoidlad ja jõed on väga bakteririkkad. Kõige rohkem baktereid leidub vee pinnakihtides, rannikule lähemal. Rannikust kaugenedes ja sügavuse kasvades bakterite arv väheneb.
Puhas vesi sisaldab 100 ml baktereid 1 ml kohta ja saastunud vesi-100–300 tuhat ja rohkem. Põhjasettes on palju baktereid, eriti pinnakihis, kus bakterid moodustavad kile. See kile sisaldab palju väävli- ja raudbaktereid, mis oksüdeerivad vesiniksulfiidi väävelhappeks ja takistavad seeläbi kalade hukkumist. Muda sisaldab rohkem eoseid kandvaid vorme, samas kui vees valitsevad eoseid mittekandvad vormid.
Liigilise koostise poolest sarnaneb vee mikrofloora mulla mikroflooraga, kuid on ka spetsiifilisi vorme. Hävitades mitmesuguseid vette sattunud jäätmeid, viivad mikroorganismid järk-järgult läbi nn vee bioloogilise puhastamise.
Õhu mikrofloora on vähem rikkalik kui pinnase ja vee mikrofloora. Bakterid tõusevad tolmuga õhku, võivad seal mõnda aega viibida ja seejärel maapinnale settida ning toitumise puudumise või ultraviolettkiirte mõjul surra. Mikroorganismide arv õhus sõltub geograafilisest tsoonist, maastikust, aastaajast, tolmureostusest jne. Iga tolmukübar on mikroorganismide kandja. Enamik baktereid on õhus tööstusettevõtete kohal. Maal on õhk puhtam. Puhtaim õhk metsade, mägede ja lumistes kohtades. Õhu ülemised kihid sisaldavad vähem mikroobe. Õhu mikrofloora sisaldab palju pigmenteerunud ja eoseid sisaldavaid baktereid, mis on teistest vastupidavamad ultraviolettkiirgusele.
Inimkeha, isegi täiesti terve, on alati mikrofloora kandja. Kui inimese keha puutub kokku õhu ja pinnasega, asetuvad riietele ja nahale mitmesugused mikroorganismid, sealhulgas patogeensed (teetanusepulgad, gaasgangreen jne). Kõige sagedamini on inimkeha avatud osad saastunud. Escherichia coli, stafülokokke leidub kätel. Suuõõnes on üle 100 tüüpi mikroobide. Suu oma temperatuuri, niiskuse ja toitainete jääkidega on suurepärane keskkond mikroorganismide arenguks.
Maol on happeline reaktsioon, nii et suurem osa selles sisalduvatest mikroorganismidest sureb. Alustades peensoolest, muutub reaktsioon aluseliseks, s.t. mikroobidele sõbralik. Käärsooles on mikrofloora väga mitmekesine. Iga täiskasvanu eritab iga päev umbes 18 miljardit bakterit, s.t. rohkem inimesi kui inimesi maailmas.
Väliskeskkonnaga (aju, süda, maks, kusepõis jne) mitteseotud siseorganid on tavaliselt mikroobivabad. Mikroobid sisenevad nendesse elunditesse ainult haiguse ajal.
Mikroorganismid üldiselt ja eriti bakterid mängivad olulist rolli bioloogiliselt olulistes aineringetes Maal, viies läbi keemilisi muundumisi, mis ei ole taimedele ega loomadele täiesti kättesaamatud. Elementide tsükli erinevaid etappe viivad läbi erinevat tüüpi organismid. Iga üksiku organismirühma olemasolu sõltub elementide keemilisest muundamisest, mida viivad läbi teised rühmad.
Lämmastikku sisaldavate ühendite tsüklilisel muundamisel on esmane roll vajalike lämmastikuvormide varustamisel biosfääri organismidega erinevate toitumisvajaduste jaoks. Üle 90% kogu lämmastiku sidumisest on tingitud teatud bakterite metaboolsest aktiivsusest.
Orgaanilise süsiniku bioloogiline muundamine süsinikdioksiidiks, millega kaasneb molekulaarse hapniku vähenemine, nõuab erinevate mikroorganismide ühist metaboolset aktiivsust. Paljud aeroobsed bakterid viivad läbi orgaanilise aine täieliku oksüdatsiooni. Aeroobsetes tingimustes lagunevad orgaanilised ühendid algselt kääritamise teel ja käärimise orgaanilised lõppsaadused oksüdeeritakse anaeroobse hingamise tagajärjel edasi, kui on olemas anorgaanilisi vesiniku vastuvõtjaid (nitraat, sulfaat või CO 2).
Väävel on elusorganismidele saadaval peamiselt lahustuvate sulfaatide või redutseeritud orgaaniliste väävliühendite kujul.
Mõned mageveekogud sisaldavad suurtes kontsentratsioonides vähendatud rauasoolasid. Sellistes kohtades areneb spetsiifiline bakteriaalne mikrofloora - raudbakterid, mis oksüdeerivad redutseeritud rauda. Nad osalevad rabamaakide ja raua soolade rikaste veeallikate moodustamises.
Bakterid on vanimad organismid, mis ilmusid Arheanides umbes 3,5 miljardit aastat tagasi. Umbes 2,5 miljardit aastat domineerisid nad Maal, moodustades biosfääri, osaledes hapniku atmosfääri moodustamises.
Bakterid on üks lihtsamaid elusorganisme (peale viiruste). Arvatakse, et need on esimesed organismid, kes Maal ilmusid.
5 MIKROORGANISMIDE SALLIMINE KESKKONNAFAKTORITELE
Mikroorganismide areng ja elutegevus on tihedalt seotud keskkonnaga. Nende tegevuse avaldumine sõltub selle keskkonna muutustest või omadustest.
Iga mikroorganismitüüp on võimeline kasvama, arenema ja paljunema välistingimuste raames, mis peegeldavad nende taluvust.
Keskkonnategurid on arvukad ja mitmekesised. Tavaliselt jagunevad need füüsikalisteks, keemilisteks ja bioloogilisteks.
Mikroorganismid kohanevad äärmuslike füüsikaliste ja keemiliste keskkonnateguritega paremini kui loomad ja taimed. Mõned bakterid jäävad elujõuliseks temperatuuril kuni + 104 ° C, pH vahemikus 1 kuni 13, rõhul 0 kuni 1400 atm. , raskmetallide, antiseptikumide, antibiootikumide, desinfektsioonivahendite juuresolekul. Samal ajal on iga liigi puhul pärilikult määratud optimaalsed tasemed ja kriitilised piirid mikroobide taluvusele füüsikaliste, keemiliste ja bioloogiliste tegurite suhtes.
Tolerantsus keskkonna füüsiliste tegurite suhtes
Väliskeskkonna füüsikalised tegurid, mis mõjutavad positiivselt või negatiivselt mikroorganismide elutegevust, hõlmavad järgmist: keskkonna niiskus, selles lahustunud ainete kontsentratsioon ja selle osmootne rõhk, temperatuur, päikesevalgus ja erinevad kiirgusenergia vormid.
Keskkonna niiskus. Teatud tüüpi mikroobid on niiskuse puudumise suhtes väga tundlikud. Näiteks surevad pärast kuivatamist kiiresti ära nitrifitseerivad ja äädikhappebakterid. Teised, vastupidi, võivad kuivaks jääda mitu kuud ja isegi aastaid (stafülokokid, piimhappebakterid, pärm). Bakterite ja hallitusseente eosed on eriti vastupidavad kuivatamisele. Neid saab aastakümneid kuivana hoida. Kuivatamine vaakumis madalal temperatuuril ja sellele järgnev õhuvabas keskkonnas säilitamine säilitab mikroobide elulise aktiivsuse pikka aega (lüofiilne). Seda meetodit kasutatakse laialdaselt mikroobikultuuride pikaajaliseks säilitamiseks. Niisiis, mõned haigusi põhjustavad bakterid (kookid) elasid sarnastes tingimustes 25 aastat ja mükobakterid 17 aastat.
Mullas arenevad mitmesugused mikroorganismide rühmad kõige intensiivsemalt niiskusesisalduse juures, mis on ligikaudu 60% kogu niiskusesisaldusest.
Kõige niiskust armastavad mullabakterid on lämmastikku siduvad bakterid (asotobakter ja sõlme). Kui pinnas kuivab, väheneb mikrobioloogiline aktiivsus või see on täielikult pärsitud. Mikroorganismide võimetust areneda ebapiisava niiskuse tingimustes kasutatakse toidu ja sööda kaitsmiseks riknemise eest kuivatamise teel. Kuivatatakse liha, kala, köögivilju, puuvilju, piima ja muid tooteid, samuti heina.
Söötmes lahustunud ainete kontsentratsioon. Looduslikes tingimustes elavad mikroorganismid lahustites, milles on erineva kontsentratsiooniga lahustunud aineid ja seega ebavõrdse osmootse rõhuga.
Soolade kontsentratsiooni suurenemine keskkonnas üle optimaalse häirib normaalset ainevahetust raku ja väliskeskkonna vahel. Sellisel juhul lahkub vesi rakust, tsütoplasma lahkub rakumembraanist (plasmolüüs) ja raku toitainete tarnimine peatatakse. Selles olekus surevad mikroorganismid üsna kiiresti ja ainult vähesed suudavad pikka aega püsida. Niisiis, on baktereid, mis on kohanenud kõrge soolakontsentratsiooniga (umbes 29%). Neid baktereid nimetatakse halofiilne("Armastav" sool).
Soolade kõrge kontsentratsiooni hävitav mõju mikroorganismidele on leidnud rakendust ka inimpraktikas. See on aluseks paljude toiduainete (liha, kala) eel serveerimiseks tugevates soolalahustes. Enamik mädanenud baktereid lakkab arenema juba 5-10% NaCl kontsentratsioonil keskkonnas (Proteus vulgaris, Bacillus mesentericus). Usaldusväärsemate tulemuste saamiseks kasutatakse siiski kontsentreeritumaid naatriumkloriidi lahuseid - 20-30%.
Kõrge osmootse rõhu tekitamiseks vedelikus kasutatakse lisaks naatriumkloriidile laialdaselt suhkruid, kuid nende kontsentratsioon ületab 70%.
Temperatuur. Keskkonna temperatuur on üks olulisemaid mikroobide elu mõjutavaid keskkonnategureid. Igat tüüpi mikroorganismid võivad areneda ainult teatud temperatuurivahemikus.
Seoses temperatuuriga jagatakse mikroorganismid tavaliselt kolme rühma: psühhofiilsed, mesofiilsed ja termofiilsed.
TO psühhofiilne(Kreeka psühhrio - külm, phileo - armastus) hõlmavad mikroorganisme, mis on madalal temperatuuril arenenud. Need on hallitusseened, helendavad bakterid, külmaveekogude bakterid, liustikud jne. Nende jaoks on minimaalne temperatuur 0 kuni 10 ° С, optimaalne umbes 10 ° С ja maksimaalne temperatuur 20-30 ° С. Mõned liigid on võimelised kasvama isegi temperatuuril alla 0 ° C.
Üldiselt on mikroorganismid madalate temperatuuride suhtes tundetud. Mitmed teadlased on näidanud, et bakterid säilitavad oma elujõulisuse pärast seda, kui neid on mitu tundi töödeldud vedela õhuga (-182,-100 ° C) või isegi vedela vesinikuga (-252 ° C). Madalad temperatuurid peatavad mikroorganismide eluea, takistades seetõttu jahutatud liha, kala, või, piima ja muude toodete riknemist. Taaskülmutamine pärast sulatamist mõjub mikroobidele kahjulikult. Psührofiilsed bakterid ei moodusta eoseid.
Mesofiilsed bakterid(Kreeka keeles inesos - keskmine) arenevad keskmisel temperatuuril. Nende hulka kuuluvad enamik saprofüüte ja kõik patogeensed mikroobid.
Nende jaoks on minimaalne temperatuur vahemikus 0-10 ° C, optimaalne temperatuur 25-35 ° C ja maksimum 40-50 ° C.
Termofiilsed bakterid(Kreeka termos-soe) areneb suhteliselt kõrgel temperatuuril. Nende minimaalne temperatuur on umbes 30 ° C, optimaalne on 50-60 ° C, maksimaalne on 70-80 ° C.
Termofiilsed mikroorganismid on kuumas mineraalvees laialt levinud ja osalevad aktiivselt sõnniku, silo ja märja teravilja isekuumenemise protsessides.
Kõrge temperatuur põhjustab mikroobrakkude surma tsütoplasmaatiliste valkude hüübimise (hüübimise) ja ensüümide inaktiveerimise tagajärjel. Enamik eoseid mittekandvaid baktereid sureb, kui neid kuumutatakse temperatuuril 60–70 ° C 15–30 minutit ja kuumutamisel temperatuurini 80–100 ° C mõneks sekundiks kuni 1–3 minutiks. Niiskes keskkonnas surevad bakterid kõrgel temperatuuril kiiremini kui kuivas, kuna aur soodustab valgu kiiret hüübimist. Paljude bakterite eosed taluvad kuumutamist kuni 100 ° C mitu tundi. Isegi kõige stabiilsemad eosed niiskes keskkonnas temperatuuril 120 ° C surevad 20-30 minuti pärast ja kuiva kuumusega (160-170 ° C) kokku puutudes-1-2 tunni pärast.
Kaks meetodit bakterite hävitamiseks põhinevad kõrge temperatuuri hävitaval mõjul: pastöriseerimine ja steriliseerimine.
Pastöriseerimise ajal kuumutatakse vedelikku temperatuuril 60–70 ° C 20–30 minutit või temperatuuril 70–80 ° C 6–10 minutit, samal ajal kui ainult bakterite vegetatiivsed vormid surevad. Pastöriseerimist kasutatakse peamiselt piima, veini, kaaviari, puuviljamahlade ja mõnede muude toodete säilitamiseks.
Steriliseerimine tähendab objekti või aine vabastamist kõigist elusolenditest. See saavutatakse 100–130 ° C kuumutamisega 20–40 minutit.
Valguse mõju. Otsene päikesevalgus tapab peaaegu igat tüüpi baktereid, välja arvatud lillad ja fotobakterid. Otsese päikesevalguse mõjul surevad bakterid mõne minuti või tunni jooksul.
Päikesevalguse bioloogiline mõju mikroelementidele tuleneb selles sisalduvatest ultraviolettkiirtest. Pärast rakku sisenemist põhjustavad need, olles adsorbeeritud elutähtsate osade, valkude ja nukleiinhapetega, fotokeemilisi ja oksüdatiivseid protsesse, millel on kahjulik mõju mikroorganismidele. Ultraviolettkiired tapavad mõne minuti jooksul vegetatiivsed vormid ja eosed.
Bioloogiliselt on kõige huvitavamad ultraviolettkiired lainepikkusega 280 kuni 230 nm. Neil on väljendunud bakteriostaatiline ja bakteritsiidne toime. Sõltuvalt kiirgusdoosist ja mikroorganismi tüübist võib ultraviolettkiirte mõju olla surmav või mutageenne.
Ultraviolettkiirgust kiirgavaid lampe lainepikkusega 254 nm kasutatakse laialdaselt nõude steriliseerimiseks, haiglate ja operatsioonisaalide õhu desinfitseerimiseks, koolides, võitluses teraviljade vastu. Ultraviolettkiirt kasutatakse ka vee, piima, kõrgel temperatuuril hävitatud materjalide steriliseerimiseks.
Kiirguse, röntgenikiirte ja elektri mõju. Raadium ja röntgenikiirgus väikestes annustes ja lühiajalise toimega stimuleerivad mõnede mikroobide paljunemist, suurtes annustes nad aga tapavad. Kõrgsageduslik elektrivool põhjustab mikroorganismide surma. Eriti tugevat mõju avaldavad neile ülikõrge sagedusega voolud.
Mehaaniliste löökide ja kõrge rõhu mõju. Mehaanilised mõjud (tugevad ja sagedased löögid) hävitavad enamiku mikroobidest. Aknaluugis liiva või klaashelmestega raputamine vähendab oluliselt elujõuliste bakterite arvu. Veekogude isepuhastumine mikroorganismidest on osaliselt tingitud vee liikumisest jõgedes ja ojades. Kõrge rõhk mõjutab mikroorganisme vähe, teatud tüüpi bakterid võivad normaalselt elada ja paljuneda meredes 9 km sügavusel, kus rõhk ulatub 9 × 10 4 kPa -ni. Teatud tüüpi pärm, hallitusseened ja bakterid taluvad survet ja 3 × 105 kPa.
Tolerantsus keskkonna keemiliste tegurite suhtes
Mikroorganismide elutegevust mõjutavad keemilised tegurid on järgmised: keskkonna koostis ja reaktsioon, keskkonna redokstingimused.
Keskkonna koostis. Keemilised ühendid võivad olla mikroorganismidele kasulikud ja toitainetena või ebasoodsad - antimikroobsed (bakteritsiidsed), mis pärsivad või tapavad mikroorganisme. Nõrgad lahendused suurendavad mikroobide elutegevust. Tugevamad lahused tapavad mikroorganisme ainult vegetatiivses faasis, väga kontsentreeritud lahused hävitavad ka eoseid. Erinevate mikroobide tundlikkus sama keemilise ühendi suhtes ei ole sama. Mõned ained mõjuvad kahjulikult mõnele mikroorganismide rühmale ja on teistele kahjutud.
Anorgaanilistest ainetest on mikroorganismidele kõige mürgisemad raskmetallide (elavhõbe, vask, hõbe) soolad. Kontsentratsioonil 1: 1000 tapetakse enamik baktereid mõne minuti jooksul. Kloor, jood, vesinikperoksiid, kaaliumpermanganaat omavad bakteritsiidset toimet. Mineraalhapetest omavad neid omadusi väävel-, boor- ja mõned muud happed.
Tugevad mürgid mikroobidele on fenool (karboolhape), kreosool, formaliin. Alkoholid ja mõned orgaanilised happed (salitsüül-, või-, äädik-, bensoehape) on erineval määral mürgised.
Liha ja kala suitsetamise aluseks on antiseptikumide hävitav toime bakteritele, mille käigus toode immutatakse lenduvaid ühendeid sisaldava suitsuga, eelkõige formaldehüüdi, fenoolide, vaikudega.
Keskkonna reaktsioon. Keskkonna reaktsioon on oluline keemiline tegur, mis mõjutab mikroorganismide elutegevust. Neutraalse keskkonna pH väärtus on 7,0, happelise keskkonna puhul - 0-6,0 ja aluselise - 8,0-14,0. Mikroobide suhtumine keskkonna reaktsiooni on väga mitmekesine. Kui mõned võivad areneda laias pH väärtuste vahemikus, siis teiste mikroorganismide arenguks peaksid pH kõikumised olema ebaolulised.
Paljude hallitusseente ja pärmide puhul on kõige soodsam keskkond, mille pH on 3,0-6,0; enamik baktereid areneb paremini neutraalses või kergelt aluselises keskkonnas (7,0-7,5). Väga happeline reaktsioon bakteritele on hävitav.
Erandiks on bakterid, mis moodustavad ise hapet (äädikhape, piimhape, liitiummonohape ja võihape).
Mullas või veekogudes elavad mikroorganismid puutuvad kokku oluliste pH kõikumistega, seega on nad kohanenud paljude pH väärtustega. Seevastu inimese või looma kehas elavad patogeensed mikroorganismid võivad areneda suhteliselt kitsas pH vahemikus.
Keskkonna redokstingimused. Mikroorganismide areng on tihedalt seotud keskkonna redokstingimustega, mida tavapäraselt tähistatakse sümboliga rH 2 . See on vesiniku molekulaarse rõhu negatiivne logaritm ja väljendab keskkonna aeroobsuse astet. Kui sööde on küllastunud molekulaarse vesinikuga, siis rH 2 on võrdne nulliga. Oksüdeerimis- ja redutseerimisprotsesside tasakaalus keskkonnas rH 2 on võrdne 28. Kui sööde on hapnikuga küllastunud rH 2 võrdne 41. Keskkonna redokspotentsiaali mõjutab õhutamine. Erinevatel mikroorganismidel on redokstingimuste kardinaalsed punktid - minimaalne optimaalne ja maksimaalne, mis määravad nende arengu.
Mikroorganismide hapnikutarve on väga erinev. Anaeroobid suudavad paljuneda madalate väärtuste korral rH 2 - 8 kuni 10. Aeroobid paljunevad vahemikus rH 2 vahemikus 10 kuni 30. Vahevormid (fakultatiivsed anaeroobid) võivad areneda laias vahemikus rH 2 - 0 kuni 30.
Reguleerides redokstingimusi keskkonnas, on võimalik mitte ainult mõjutada mikroorganismide kasvu ja arengut, vaid ka mõjutada mikroorganismide poolt põhjustatud füsioloogiliste ja biokeemiliste protsesside olemust.
Tolerantsus bioloogiliste keskkonnategurite suhtes
Mikroobide suhete tüübid biotsenoosides.
Mikroorganismid on karmid võistlema omavahel. See on tingitud asjaolust, et teatud biotsenoosis elavatel mikroobidel on põhimõtteliselt sarnased energia- ja toitumisallikate vajadused. Iga mikroorganism kohaneb mitte ainult elutute substraatidega, vaid ka teiste ümbritsevate organismidega. Selline kohanemine viib mõnikord spetsiaalsete metaboolsete omaduste omandamiseni, mis annavad omanikule võimaluse hõivata konkreetseid nišše. Näiteks võivad nitrifitseerivad bakterid valguse puudumisel kasvada ilma orgaaniliste energiaallikateta, oksüdeeriva ammoniaagi või nitritita energiaallikana; teised organismid ei arene sarnastes tingimustes. Seetõttu pole nitrifitseerivatel bakteritel bioloogilist konkurentsi. Märkimisväärne osa baktereid osaleb konkurentsivõitluses, kohanedes kooselu teiste eluvormidega või astudes nendega opositsiooni.
Sümbioos. Sümbioosi näiteks on seos mõne piimhappebakteri ja pärmi vahel (piimhappebakterid loovad piimhapet tootes pärmi kasvuks soodsad tingimused ja pärmijäätmed - vitamiinid stimuleerivad piimhappebakterite arengut), lämmastikku siduvad mikroobid ja tselluloosi lagundavad bakterid aeroobid, absorbeerivad hapnikku, anaeroobidega jne. Sellist seost täheldatakse sageli mikroorganismide ja taimede vahel (näiteks mügarbakterite sümbioos kaunviljadega, mükoriisa on erinevate taimede juurtega seente kooselu), samuti mikroobide ja loomade vahel.
Suhe, milles mikroorganism asub väljaspool peremehe (suurema organismi) rakke, on tuntud kui ektosümbioos; kui lokaliseeritakse rakkude sees - nagu endosümbioos.
Tüüpilised ektosümbiootilised mikroobid - Escherichia coli, perekonna bakterid Bakteroidid ja Bifidobakterid, Proteus vulgaris, samuti teised soole mikrofloora esindajad.
Sümbiootilisel suhtel on järgmised vormid.
Metabioos - selline olemasolu, kui teatud tüüpi mikroobide jääkained on materjalid teiste liikide toitumiseks ja arenguks. Näiteks lagundavad saprofüüdid looduslikud valgud peptionideks, aminohapeteks ja muudeks lihtsamateks ühenditeks. Ja need tooted on lähteaineks nitrifitseerivatele bakteritele, mis muudavad ammoniaagi soolad lämmastikuks ja seejärel lämmastikhappeks.
Pärm muudab suhkrud etüülalkoholiks ja äädikhappebakterid oksüdeerivad selle äädikhappeks. See suhtlusvorm on levinud mulla mikroobide seas ja on looduses ainete ringluse aluseks.
Kommensalism(lat. com + mensa - kaaslased) - omamoodi sümbioos, millest saab kasu ainult üks partner (tekitamata teisele nähtavat kahju). Mikroorganismid-kommensaalid koloniseerivad inimkeha nahka ja õõnsusi (näiteks seedetrakti), põhjustamata "nähtavat" kahju; nende kogu on normaalne mikroobne taimestik (looduslik mikrofloora). Tüüpilised ektosümbiootilised kommensaalsed organismid on Escherichia coli, bifidobakterid, stafülokokid, laktobatsillid. Paljud kommensaalbakterid kuuluvad oportunistlikku mikrofloorasse ja võivad teatud tingimustel põhjustada makroorganismi haigusi (näiteks kui need viiakse meditsiiniliste protseduuride käigus vereringesse).
Vastastikkus (lad. mutuus - vastastikune) - vastastikku kasulik sümbiootiline suhe. Seega toodavad mikroorganismid peremeesorganismi jaoks vajalikke BAB -sid (näiteks B -vitamiine). Samal ajal on makroorganismides elavad endo- ja ektosümbiontid kaitstud ebasoodsate keskkonnatingimuste (kuivamise ja äärmuslike temperatuuride) eest ning neil on pidev juurdepääs toitainetele. Igasugusest vastastikkusest on kõige üllatavam mõnede seente kasvatamine putukate (mardikad ja termiidid) poolt. Ühest küljest aitab see kaasa seente laiemale levikule, teisest küljest pakub see vastsetele pidevat toitainete allikat.
Satelliit. Mõned mikroorganismid on võimelised eritama metaboliite, mis stimuleerivad teiste mikroorganismide kasvu. Näiteks sarkiinid või stafülokokid eritavad kasvufaktoreid, mis stimuleerivad perekonna bakterite kasvu Haemophilus... Sageli aktiveerib mitut tüüpi mikroobide ühine kasv nende füsioloogilisi omadusi. Selliseid suhteid tuntakse kui satelliitsmi (ladina keelest safeties - kaasas) (joonis 6).
Riis. 6. Sünergism mikroobides - agariploki ümber aktinomütseetide kultuuriga on nähtav hallitusseente kasvu stimuleeriv tsoon.
Antagonism (antibioos) - Olukordi, kus üks mikroorganism pärsib teise arengut, nimetatakse mikroobide antagonismiks (kreeka keelest antagonizmai - rivaalitsemine) ja need peegeldavad mikroorganismide võitlust evolutsiooni nimel (see tähendab toidu- ja energiaallikate pärast).
Antagonistlikud suhted on eriti väljendunud paljude erinevate mikroorganismide liikide ja liikide looduslikes elupaikades (näiteks mullas või seedetraktis), millel on samad toitumis- ja energiavajadused. Sellisel juhul võib mõju konkurendile olla passiivne või aktiivne. Esimesel juhul kasutavad mikroorganismid substraati kiiremini ära, jättes rivaali "toorainest" ilma; teises - "kuulutada sõda kuni täieliku hävinguni". Hävitamise vormid võivad olla erinevad - alates väiksemate liikide banaalsest imendumisest kuni väga spetsiifiliste toodete eraldamiseni, mis on konkurentidele mürgised (joonis 7).

Riis. 7. Antagonism mikroobides - stafülokokkide kasvu pärssimise tsoon on nähtav agarinploki ümber koos aktinomütseekultuuriga.
Antagonistlike mikroorganismide hävitav toime on seotud jäätmete kogunemisega keskkonda või teatud bioloogiliselt aktiivsete ainete - antibiootikumide - vabanemisega sellesse.
Sellise kahjuliku mõju tagajärjel nõrgeneb ühe liigi elutegevus või ta sureb.
Piimhappebakterid on mädanenud bakterite antagonistid, kuna piimhape pärsib nende arengut. Tavaline mulla mikrofloora pärsib mikroorganisme, mis on inimestele patogeensed.
Antagonismi täheldatakse ka taimede ja mikroorganismide vahel. Taimed toodavad aineid, mis on mürgised bakteritele, seentele ja algloomadele. Nendel ainetel on erinevad omadused ja need ei ole ühesugused keemilise olemuse, toime tugevuse jms poolest. Esmakordselt tuvastas need 1928. aastal Nõukogude botaanik V. P. Tokin ja nimetas need fütontsiidid(füton - taim, kaedo - tappa).
Seega on mikroobimaailma taluvuspiirkond tõeliselt tohutu, selle piirid on sageli keskkonnategurite äärmuslikel väärtustel. See mikroorganismide omadus tagab neile peaaegu piiramatu arengu kogu planeedil.
RAKENDUSBIOKEEMIA JA MIKROBIOLOOGIA, 2004, köide 40, nr 4, lk. 387-397
UDC: 576.8.098 / 577.1
BAKTERITE KOHALDAMISEKS KESKKONNATINGIMUSTEGA VÄLISED TEGURID
© 2004 Yu. A. Nikolaev
Mikrobioloogia Instituut RAS, 117811, Moskva, e-post: [e -post kaitstud] Saadud 17. novembril 2003
Arvesse võetakse andmeid bakterite rakuväliste ühendite kohta, mis on seotud nende kohanemisega ebasoodsate keskkonnatingimustega: kõrged ja madalad temperatuurid, kasvu pärssivad ja bakteritsiidsed toksiliste ainete (oksüdeerijad, fenoolid, raskmetallid) kontsentratsioonid, antibiootikumid, ebasoodne pH ja soolsus. Tuvastatud ühendid on oma olemuselt erinevat tüüpi; neid esindavad valgud, süsivesinikud, orgaanilised happed, nukleotiidid, aminohapped, lipopeptiidid, lenduvad ühendid. Enamikku neist ühenditest pole veel kindlaks tehtud ja nende omadusi on uuritud biotestide abil. Tehakse ettepanek kaaluda rakuväliseid kohanemisfaktoreid (EFA) kui uut bioloogiliselt aktiivsete ainete rühma. Toimemehhanismi järgi saab rakuväliseid kohanemisfaktoreid jagada mitmeks rühmaks; kaitsmed (stabilisaatorid); signaallaadi ained, mis on rakkude kaitsemehhanismide indutseerijad; regulaatorid ei ole induktiivpoolid (näiteks adhesiooniregulaatorid); "vastumürk" ja neutraliseeriv toime. HFA uuringu peamised suunad on uute ühendite otsimine (biotestide põhjal), nende tuvastamine ja toimemehhanismide uurimine. Rakuvälised kohanemisfaktorid võivad leida laialdast praktilist rakendust biotehnoloogias, meditsiinis, põllumajanduses ja keskkonnakaitses.
Bakterite kohanemine ebasoodsate keskkonnatingimustega on traditsiooniline ja hästi uuritud biokeemia ja mikrobioloogia haru. Kohanemist (ladina ayargagu - kohanemine) mõistetakse kui keha füsioloogiliste, biokeemiliste, morfoloogiliste ja käitumuslike reaktsioonide summat, mille eesmärk on muuta kasvukiirust, ainevahetust, elujõulisust (ellujäämist) ja kehale omane. Kohanemine on suunatud konkreetse populatsiooni ja kogu liigi kui terviku ellujäämisele. Mikrobioloogia, biokeemia ja teoreetilise bioloogia õpikutes käsitletakse kohanemist ebasoodsate keskkonnatingimustega rubriikides "fenotüüpne ja geneetiline kohanemine" ning "ensüümide aktiivsuse ja nende sünteesi reguleerimine". Kohanemise spetsiifilised näited on mitmekesised ning neid on kirjeldatud mitmes arvustuses ja monograafias, peamiselt biokeemia ja adaptiivse reaktsiooni arengu geneetilise kontrolli seisukohast.
Peatume lühidalt terminoloogial, sest isegi selles valdkonnas töötavate spetsialistide seas puudub spetsiifiliste terminite kasutamisel ühtsus. Ingliskeelses kirjanduses räägivad nad tavaliselt kohanemisest kui stressile või šokkidele (hape, temperatuur, sool jne) vastupidavuse kujunemisest, stressi mõistmisest rõhuna, pingena, rõhuna, teisisõnu - mõnel juhul oluline muutus. tegur - temperatuur, rõhk jne .NS. "Šokk"
tähendab lööki, šokki, tõukamist, s.t. terav mõju kehale, lühiajaline võrreldes rakutsükli pikkuse ja normaalsete kohanemisreaktsioonide kiirusega ning märkimisväärne teguri intensiivsuse poolest.
Vene bioloogias on erinev terminoloogia. Vene keele seletava sõnaraamatu järgi on "stress kehas pingeseisund, kaitsereaktsioon, mis on põhjustatud ebasoodsa teguri toimest". See arusaam on kooskõlas stressiteaduse rajaja G. Selye määratlusega, kes tõlgendas stressi kui keha mittespetsiifilist vastust talle esitatud nõudmisele. Stressi käsitletakse mitmes arvustuses samas tähenduses. Šokk on "keha reaktsioon tugevale välismõjule (samuti selle seisund pärast sellist lööki), mida iseloomustab eluprotsesside reguleerimise järsk rikkumine". Seega on nii stress kui ka šokk keha seisund, iseloomulik adaptiivne reaktsioon mõjudele, mis erinevad doosi, intensiivsuse ja aja poolest.
On õigustatud käsitlema küsimust, kui loomulik, tavaline või ebanormaalne on stress. Sel juhul tuleks märkida, mida peetakse normiks. Pidevad muutused, areng on bioloogiliste süsteemide lahutamatud omadused.
Muutused on nii vektor-, ühesuunalised kui ka tsüklilised. Sellest tulenevalt peaksid iga elusüsteemi arendamisel vahelduma kiire, optimaalse arengu faasid ja allasurutud, mitteoptimaalse arengu faasid. Selle põhjal tuleks allasurutud, piiratud olekuid pidada elu loomulikuks ja isegi lahutamatuks omaduseks. Sellisel juhul, kui mikroorganismide paljunemist, nende arvu suurenemist mikroobikultuuri kasvufaasis, peetakse normiks, siis loetakse rakkude seisundit mahajäämuse faasis uue stressiks. keskkond ”, mis nõuab adaptiivseid vastuseid. Teisest küljest põhjustab toiduallikate ammendumine või rakkude tiheduse kriitiline suurenemine arenevas kultuuris "näljastressi" ja statsionaarsed rakud näitavad kultuuri füsioloogilist kohanemist tingimustega, mis ei ole kasvamiseks optimaalsed. Selline stressi käsitlemine normaalse ja isegi kultuuriseisundi (organismi) arengut soodustava tegurina on seotud ja kombineeritud selle konkreetse füüsilise maailma tsüklilisusega, kus maismaaorganismid eksisteerivad, kus temperatuur, valgustus, niiskus, rõhk, orgaaniliste ja anorgaaniliste ühendite kontsentratsioonid, füüsiline pinge muutub tsükliliselt, väljad, biootiliste tegurite toime.
Võttes arvesse keskkonnatingimuste muutuste tsüklilisust, tuleks neis eristada: a) muutusi, mis on teatud arenguetapis uued, kuid arengutsüklis korduvad ja kuuluvad antud liigi taluvustsooni; b) mõju väljaspool sellist taluvuspiirkonda, mis on organismi kasvule ja arengule ebasoodne, sageli biotsiidne. Seejärel peame igasuguste tingimustega kohanemise all silmas keha spetsiifiliste morfoloogiliste, biokeemiliste, füsioloogiliste ja käitumuslike reaktsioonide kogumit, mis arenevad vastuseks nendele tingimustele ja aitavad kaasa keha toimimise jätkamisele või mille eesmärk on suurendada elujõudu (vähendada suremust) ekstreemsed tingimused. Viimasel juhul ei räägi me ainevahetuse (mikroobikultuuri) jätkumisest konkreetsetes ebasoodsates tingimustes (organismi kohanemine), vaid populatsiooni säilimisest järgmiseks elutsükliks kauges tulevikus koos surmaga. osa elanikkonnast (tavaliselt suurem) ja ellujäänute rakkude toimimise ajutine seiskumine.
Arutatava probleemi kontekstis peatume lühidalt mikroorganismide stressimärkidel. Kui see on kindlaks tehtud, on vaja võrrelda organismi seisundi näitajaid, mida täheldatakse tavalises, optimaalseks (homöostaas) ja stressiseisundis.
nii. Homöostaasi mis tahes mõõdetava näitaja oluline kahjustus on stressi näitaja ja märk. Mikroorganismides hõlmavad depressiivse ja stressiseisundi näitajad järgmist: valgu vabanemine rakuvälisesse ruumi, väikese molekulmassiga ühendite kadumine raku poolt tsütoplasmaatilise membraani suurenenud läbilaskvuse tõttu, ribosoomide, nukleiinhapete kahjustus, kiiruse vähenemine. hapniku tarbimine, ensüümide aktiivsuse vähenemine, reaktiivsete hapnikuühendite ja lipiidide peroksüdatsiooniproduktide kuhjumine, osa elanikkonna rakkude võime kaotada kolooniad kasvamise ajal tihedas toitainesisalduses (st. (kolooniaid moodustavate üksuste kontsentratsioon, CFU), kasvupeetus, elutähtsa aktiivsuse pärssimine, agregatsioon ja adhesioon. Kohustuslik märk stressirohke olekust on selle pöörduvus, võime naasta normaalse toimimise juurde koos sobivate keskkonnamuutustega.
Loetletud märkide hulgas on kõige iseloomulikum ja praktikas kõige sagedamini kasutatav kaks - kasvukiiruse ja rakkude elujõulisuse vähenemine. Need tunduvad olevat kõige otsesemad ja adekvaatsemad stressiseisundi näitajad. Kasvukiirus on mikroorganismide seisundi lahutamatu näitaja. Rakkude puhul, mis kasvavad teatud kiirusel, maksimaalselt antud tingimustel (cmax), näitab selle vähenemine mõnele madalamale cmin väärtusele mikroobide stressiseisundit. Hiljem võib aga selle uue madala tempoga kasv muutuda uues keskkonnas normiks. Tingimuste edasine halvenemine võib viia q vähenemiseni 0 -ni või isegi rakusurmani. Rakusurma algus näitab üksikute rakkude adaptiivsete ressursside ammendumist. Kuid populatsiooni jaoks on rakkude arvu vähenemine pöörduv protsess ja teatud arvu üksikute rakkude surm on täiesti normaalne nähtus, hoolimata asjaolust, et teine osa rakkudest jääb elujõuliseks, muutudes puhkeolekusse . Seega on elujõuliste rakkude kasvukiiruse ja kontsentratsiooni vähenemine stressi tunnused, kuid esimene iseloomustab rohkem rakkude ja teine populatsiooni seisundit. Enamik töid, mis on pühendatud bakterite kohanemisele kahjulike mõjudega, toimivad täpselt nende kahe näitajaga.
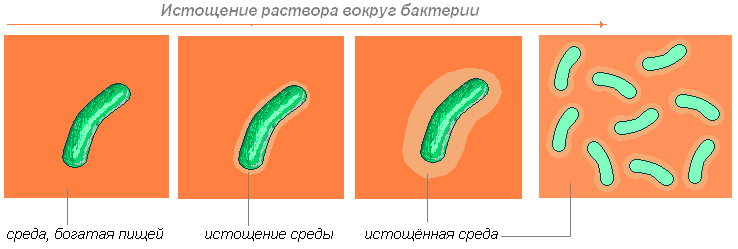
Läbivaatamist piirab peamiselt aktiivselt kasvavate kultuuride kohanemine ebasoodsate füüsikalis -keemiliste keskkonnatingimustega, mille eesmärk on nii rakkude aktiivse seisundi säilitamine, kui kasvustrateegiat säilitatakse, kui ka populatsiooni säilitamine kasvustrateegia muutmisel.
kogemuste strateegia. Mikroobikultuuride kohandamist selliste tingimustega nagu toitainete ammendumine, uued soodsad tingimused, toiduallikate muutmine, mis on sageli seotud põllukultuuride ontogeneesiga (st uinuvate vormide teke ja idanemine), ei võeta arvesse.
Praegu on hästi uuritud mehhanisme, kuidas bakterid kohaneda kõrge ja madala temperatuuriga, reaktiivsete hapnikuühendite kõrge kontsentratsiooniga, soolad, mitteioonsed ained, kõrge kiirgus, hüdrostaatiline rõhk, süsinikuallikate ammendumine, energia ja muud ressursid. Tähelepanu keskmes on rakusisene
A. V. Oleskin - 2009
Yu.V. Zaitseva, A. A. Popova ja I. A. Khmel - 2014